Продуктивность водоемов - их свойство создавать органическое вещество обычно оценивают по уровню первичной продукции планктона, рассчитываемой чаще всего за год или вегетационный сезон. Исследованиям первичной продукции планктона посвящена огромная литература. Наиболее полный ее анализ по отношению к континентальным водоемам был выполнен В.В. Бульоном, что позволило ему установить многие закономерности (Бульон, 1994).В меньшей степени известна продуктивность лотических экосистем. Однако продукционные возможности водоемов или водотоков могут быть оценены более полно с учетом продукции водорослей планктона, макрофитов, перифитона, фитобентоса, Суммарную продукцию всех автотрофов, отражающую величину первичной водоема, назовем первичной продукцией экосистемы.
Первичная продукция экосистемы в целом (Р ре) складывается из первичной продукции планктона, перифитона, макрофитов, и т.д. В различных водоемах вклад каждой из составляющих в первичную продукцию экосистемы различен (Алимов, 1989). В реках и некоторых озерах общая первичная продукция определяется в основном продукционными возможностями макрофитов и перифитона, в большинстве озер основная роль в создании первичной продукции принадлежит водорослям планктона (табл. 5).
В целом отмечается тенденция увеличения роли макрофитов и перифитона в образовании первичной продукции экосистемы в мелководных озерах. В глубоководных озерах первичная продукция создается главным образом за счет фотосинтетической активности фитопланктона. Значение перифитона среди первичных продуцентов зависит от особенностей конкретных водоемов.
Таблица 5
Доля (%) продукции водорослей фитопланктона, макрофитов, перифтона в первичной продукции водоемов и водотоков (из Function ...,1980)
|
Водоем, водоток |
глуб. сред.,м. |
Макрофиты |
Перифитон |
Фитопланктон |
|
Бере-Стрим, Англия | ||||
|
Рут-Спринг, США | ||||
|
Сильвер-Спринг, США | ||||
|
р.Темза, Англия | ||||
|
оз.Лоуренс, США | ||||
|
оз.Марион, Канада | ||||
|
оз.Боракс, США | ||||
|
оз.Латниярви, Швеция | ||||
|
оз.Миколайское, Польша | ||||
|
оз.Баторино, Белоруссия | ||||
|
оз.Нарочь, Белоруссия | ||||
|
оз.Красное, Россия | ||||
|
оз.Пяярви, Финляндия | ||||
|
оз.Субайа, Африка | ||||
|
Киевское в-ще, Украина |
Обобщение данных по продукции макрофитов (воздушно-водных и погруженных) и первичной продукции планктона позволило М.В. Мартыновой (1984) выделить в зависимости от их соотношения пять групп водоемов. Доля продукции макрофитов от суммарной первичной продукции (макрофитов и планктона) в водоемах первой группы составляла более 60, второй - 59-30, третьей - 29-11, четвертой - 5-10, пятой - менее 5 %.
Выполненные автором расчеты по данным М.В.Мартыновой (1, 2, 4 -я группы) показали, что с увеличением первичной продукции планктона возрастает продукция макрофитов(Р m), что может быть выражено в виде уравнений линейной функции (все в гС/м 2 год):
1-я группа - Р m = 1.296 Р р + 65.98 , R 2 = 0.68,
2-я группа - Р m = 1.54 Р р - 93.949, R 2 =0.83
3 -я группа - Р m = 0.26 Р р - 0.47, R 2 = 0.85 (рассчитано Мартыновой),
4-я группа - Р m = 0.117 Р р - 5.007, R 2 = 0.83,
5-я группа - Р m = 0.025 Р р + 0.31, R 2 = 0.83 (рассчитано Мартыновой).
Скорость изменения величины продукции макрофитов величины продукции планктона (первые производные приведенных уравнений) в целом снижается по направлению от 1-ой к 5 -ой группе водоемов. В тех водоемах, в которых продукция макрофитов составляет от 60 до 90% первичной продукции водоема с ростом продукции планктона наиболее резко увеличивается продукция макрофитов и, наоборот, в водоемах, где свыше 90% первичной продукции составляет продукция планктона рост продукции макрофитов происходит с малыми скоростями. При этом, по данным Мартыновой, площадь зарастания водоема макрофитами (G) нарастает пропорционально скорости увеличения соотношения между продукцией макрофитов и первичной продукцией планктона (рис.20):
G = 53.013*(d P m /d P p) 1.001 ; R 2 =0.73.(24)
В то же время площадь зарастания макрофитами резко возрастает по мере увеличения емкости водоема (Е = h/h max)(рис.21):
G = 757.67*Е 4.35 ; R 2 = 0.65 (25)
Из приведенных уравнений нетрудно видеть, что:
d P m /d P p = 8.47*Е,
т.е. продукция макрофитов по отношению к первичной продукции планктона больше в более мелководных водоемах. Обычно средняя глубина в водоемах первой группы не превышает 1-1.5 м в то время как водоемы 4-ой и особенно 5-ой групп могут иметь средние глубины 10 и более м. В первом случае макрофиты занимают почти 100% акватории, в последнем площадь зарастания макрофитами составляет доли или единицы процентов акватории.
Роль перифитона зависит от особенностей конкретных водоемов, и в некоторых озерах перифитон может создавать до 70% от общей первичной продукции. Скорость фотосинтеза водорослей перифитона в разных водоемах варьирует в широком диапазоне (из Function...,1980).
Продукция водорослей перифитона может быть значительной в литорали некоторых озер, на отдельных участках рек и ручьев, особенно в их верховьях, где водоросли перифитона могут быть единственными первичными продуцентами.
Некоторые сведения о величинах первичной продукции экосистемы в лотических экосистемах говорят о том, что первичная продукция в них ниже, чем в лимнических экосистемах (табл. 6).
Таблица 6
Величины первичной продукции (Р ре, гО 2 /м 2 сутки) в озерах и реках
|
источник |
||
|
Антарктические |
Ecosystems of the world,1984 |
|
|
Арктические |
Ecosystems of the world,1984 |
|
|
Большезем. Тундра |
Трифонова,1990 |
|
|
Латгальские |
Трифонова, 1990 |
|
|
Белорусские |
Экологическая система..,1985 |
|
|
Киннерет | ||
|
Экваториальные (Африка) |
Ecosystems of the world, 1984 |
|
|
Д. Востока |
Богатов, 1994 |
|
|
Богатов, 1994 |
||
|
реки, ручьи |
Анализ данных по 134 водоемам разных широт и континентов, о которых говорилось в предыдущей главе, позволил показать, что первичная продукция планктона в водоемах (Р р, ккал/м 2 год) возрастает по мере увеличения изменчивости температуры воды в течение года или в период открытой воды (t o). Интересно, что и у водных животных ряд продукционных показателей выше при переменных температурах воды (Галковская, Сущеня, 1978). Изученные водоемы разных широт и континентов образуют четыре группы, в пределах каждой из которых прослеживается общая закономерность: возрастание первичной продукции сопровождается увеличением изменения температуры (рис. 22), что для каждой из групп может быть описано уравнениями степенной функции:
I-я группа: Р р = 4.56t o 1.71 , R 2 = 0.64 , (26)
II-я группа: P p = 252.2 * t o 0.739 , R 2 = 0.68 (27)
III-я группа: P p = 3995*t o 0.14 , R 2 = 0.76 (28)
IY-группа: Р р = 5146.6*t o 0.25 , R 2 = 0.9. (29)
Каждая из групп водоемов может быть охарактеризована средним уровнем первичной продукции (Р р), пределами изменения температуры (t o C), географическим положением. Эти и другие характеристики приведены в таблице 7.
При этом во II-ю группу водоемов не были включены озера Исландии (65 о и 64 о с.ш.), поскольку температура воды в них отличалась от обычной для водоемов на этих широтах. Были включены водохранилища: в I-ой группе водоемов - 4, во II-ой – 4, в III-ей – 2, в IV-ой – 1. Минимальная зарегистрированная величина Рр, отмеченная в антарктическом озере Верхнее (0.58 гС/м 2 год, Kaup, 1992) не учитывалась.
Таблица 7.
Некоторые характеристики водоемов, разных групп
|
водоемов |
t o min - t o max |
Рр min - Рр max ккал/м 2 ·год |
S t min - S t max | |||||||
Примечание: 1. При расчетах средней широты места в I группу не включены Японские озера (Юноно и Татсу-кума 36 о с.ш.), расположенные на высоте около 2000 м ур.моря, другие обозначения в тексте.
Из данных таблицы 7. видно, что, как и следовало ожидать, продуктивность водоемов нарастает по направлению от арктических к тропическим.
Для количественного определения среднего уровня продуктивности, используя уравнения (26-29), рассчитаем для каждой группы среднюю скорость изменения продуктивности при изменении температуры на 1 о С. С этой целью определим первую производную для каждого из уравнений, и затем, взяв определенный интеграл по диапазону изменения температуры для каждой группы, отнесем его к этому диапазону. В результате для каждой группы водоемов получим некоторую среднюю скорость изменения первичной продукции планктона, т.е. продуктивности водоема, при изменении t o на 1 о С.
Например, для 1-ой группы(26) первая производная:
dР р / d t o = 7.94* t 0.71 (30)
Средняя скорость изменения первичной продукции планктона (U,ккал/ о С) в диапазоне t o =t o 1 - t o 2 =1.5 o - 22 o C оказывается равной:
U = [dP p /dt)dt/(t 2 - t 1) = 44.1 ккал/ о С.
Рассчитанные аналогичным образом значения U для других групп изученных водоемов приведены в табл.7.
Продукционные возможности водоемов кроме световых и температурных условий определяются так же содержанием и соотношение в воде биогенных элементов. При этом отношение N:P отражает источник биогенов. Оно высокое в олиготрофных озерах, т.к. они получают биогены из ненарушенных или слабо нарушенных водосборов, для которых характерен в большей степени экспорт азота; мезотрофные и эвтрофные водоемы получают различную смесь природных источников, что снижает соотношение между азотом и фосфором, количества азота и фосфора, получаемые с водосбора эвтрофными озерами по своим свойствам близки к таковым в сточных водах (Downing, McCauley, 1992).
Анализ данных табл. 7 показал, что скорость изменения продуктивности (U) при изменении температуры воды на 1 о С увеличивается с севера на юг и достигает наибольшей величины (311.7 ккал/ о С) в экосистемах тропических водоемов.
Наибольшая скорость приращения энергии в первичной продукции, отмечаемая в экосистемах субтропических и тропических водоемов обусловлена мало меняющимися условиями внешней среды, в частности высокими температурами воды, преобладанием содержания азота в воде, которое скорее всего связано с особенностями почв на водосборе и более высокой скоростью оборачиваемости фосфора в таких экосистемах. Косвенным тому подтверждением может служить соотношение азота и фосфора в атмосферных осадках, выпадающих на разных широтах (из Ecosystems of World, 1984):
о с.ш. 0 45 50 68 75
N: P 96 26.7 19.1 22.5 18
На общее возрастание первичной продукции планктона в водоемах по направлению от высоких широт к низким обратили внимание при анализе результатов по МБП, представив такую связь в виде прямой линии при нарастающем разбросе данных в низких широтах (Brylinsky, Mann, 1973). Позднее В.В. Бульон (1994) впервые привел кривую снижения максимальных величин первичной продукции на широтах от 40 о до 80 о с.ш. Приведенная на рис.23 кривая, огибающая наибольшие значения первичной продукции в водоемах, расположенных от 0 о до 75 о с.ш., а так же в некоторых озерах от 0.5 о до 38 о ю.ш. на большом числе исследованных водоемов описывает изменения первичной продукции планктона в экосистемах водоемов разного географического положения. При этом наибольший разброс данных отмечен так же для водоемов, расположенных на широтах близких к 10 о с.ш.
Другая важная функциональная характеристика экосистемы - количество энергии, рассеиваемой всеми организмами в процессах обмена, которое может быть рассчитано как их траты энергии на обменные процессы(R e).. Ранее (Wetzel et al., 1972; Алимов, 1987) было предложено понятие продукции экосистемы (Р е). Продукция экосистемы рассматривается как разность между первичной продукцией экосистемы (Р ре) и тратами на обмен всеми гидробионтами экосистемы (Р е = Р ре - R e).
Связь продукции экосистемы (Р е, ккал/м 2 год) и биомассы всех гидробионтов в ней за тот же период времени (В е, ккал/м 2) может быть представлена в виде степенных уравнений:
учитывается только первичная продукция планктона (рис. 24):
Р е = 2.073*В е 0.876 , R 2 = 0.761, (31)
учитывается первичная продукция планктона и макрофитов (рис.25):
Р е = 5.764*В е 0.718 , R 2 = 0.748 (31а)
(Р/В) е = 2.073*В е –0.133 и (Р/В) е = 5.764*В е –0.282 .
C учетом пределов изменений В е в изученных водоемах от 83 до 2139 в (31) и от 30 до 6616 ккал/м 2 в (31а) получаем, что в первом случае коэффициенты (Р/В) е изменяются от 1.152 до 0.748 , во втором – от 2.203 до 0.482, средние их величины составляют 0.952 и 1.346 год –1 , соответственно. Следовательно, средняя величина этого коэффициента с учетом только первичной продукции планктона не отличается от единицы, т.е. продукция водорослей планктона в водоемах оборачивается за год.
Из сказанного следует, что скорость оборота биомассы в экосистемах снижается с увеличением биомассы в экосистеме и она меньше в тех случаях, когда в расчетах продукции экосистемы принимается во внимание лишь первичная продукция планктона.
Необходимо подчеркнуть, что уравнения (31и 31а) рассчитаны на основе средних за год данных для разных водоемов, и они не могут быть, естественно распространены на сезонные или межгодовые изменения первичной продукции в одном и том же водоеме.
Важным показателем функционирования экосистемы может служить отношение общих трат на обмен гидробионтами к их суммарной биомассе (R/B) е - отношение Шрёдингера. Оно служит мерой экологического оборота энергии и его рассматривают как отношение затрат энергии на поддержание жизнедеятельности к энергии, заключенной в структуре, или меру термодинамической упорядоченности - чем больше биомасса, тем больше затраты на ее поддержание.
Анализ данных для водоемов, которые были использованы для расчетов выше приведенных уравнений, не показал какой-либо закономерности изменения отношения (R/B) e при изменении продуктивности водоемов (табл.8). С вероятностью 0.05 среднее значение этого отношения находится в переделах 6.1 - 2.99 . В водоемах разного типа и разной продуктивности траты энергии на обменные процессы у гидробионтов в среднем в 4 раза превышают их биомассу.
Таблица 8
Величины отношения (R/B) e в водоемах разной продуктивности
при Р е 0
|
Название водоема |
Р ре, какл/м 2 -год |
Примечание |
|
|
Белоруссия |
|||
|
Ленинградская обл. |
|||
|
Водохранилища: | |||
|
Иваньковское | |||
|
Веселовское | |||
|
среднее 4.34, | |||
|
= ±3.77, m=±1.14 |
В большинстве изученных водоемов разность между энергией, заключенной в первичной продукции экосистемы и рассеиваемой в процессах обмена гидробионтов отрицательна. Для таких экосистем естественно не применимо понятие продукции экосистемы. Величина отношения (R/B) e в экосистемах таких водоемов, различающихся по продуктивности, так же незакономерно меняется (табл.9).
Средняя величина этого отношения в таких экосистемах составляет 12.86 (с вероятностью 0.05 не выходит за пределы 6.5 - 19.22) и превышает в 3.4 величину этого отношения для водоемов с Р е >0. Поскольку невозможно представить себе водоем, в биотических потоках экосистемы которого не участвовали бы аллохтонные органические вещества, то можно принять, в первом приближении, что для некоторого водоема со средними для экосистемы характеристикам отношение Шрёдингера может быть принято равным (3.43 + 12.86)/2 = 8.15.
Таблица 9.
Величина отношения (R/B) e в водоемах разной продуктивности
при Р е < 0
|
название водоема |
Р ре ккал/м 2 -год |
примечание |
|
|
Белоруссия |
|||
|
Баторино | |||
|
Забайкалье |
|||
|
Цаган-Нор | |||
|
Баин-Цаган | |||
|
Баин-Булак | |||
|
Зун-Торей | |||
|
Ленинградская обл. |
|||
|
Водохранилища: | |||
|
Киевское | |||
|
Рыбинское | |||
|
Иваньковское | |||
|
Угличское | |||
|
Куйбышевское | |||
|
Горьковское | |||
|
Саратовское | |||
|
среднее 12.86 | |||
|
=±7.93, m=±2.04 |
Таким образом, затраты энергии на поддержание структуры в экосистемах водоемов, существующих главным образом за счет притока внешней энергии значительно выше, чем в тех, которые могут существовать за счет только своих продукционных возможностей. Следовательно, существование таких экосистем возможно лишь при поступлении извне значительных количеств энергии. Это может быть просто поступление аллохтонных органических веществ с водосбора или поступление биогенных элементов, приводящих к эвтрофированию, или поступление органических загрязнений, так же способствующих эвтрофикации и т.д.
Связь структурных и функциональных характеристик
Несомненно, что структура и функционирование экосистем и их составляющих должны находится в очень тесной связи, ибо именно они отражают основные свойства объекта. В качестве функциональных характеристик сообществ организмов и экосистем могут быть использованы такие как продуктивность, скорость оборота биомассы, количество диссипируемой энергии, соотношение между продукцией и диссипируемой энергией или отношение Шрёдингера. Отношение продукции и диссипируемой энергии показывает соотношение между энергией на выходе из системы и энергией, рассеиваемой организмами в процессах обмена в виде тепловой. При этом продукция сообществ животных учитывает продукцию хищных, нехищных животных и количество пищи, потребляемой хищниками внутри сообщества.
Рассмотрим связь структурных и функциональных характеристик на примере сообществ животных. Отношение продукции сообществ животных (Р b), учитывающей продукцию хищных и нехищных животных и количество пищи, потребляемой хищниками внутри сообщества к тратам животных на обменные процессы (R b) и индекс разнообразия как обобщенная характеристика структурной сложности сообщества находятся между собой в обратной зависимости (Алимов, 1989):
P b /R b = *e - H ,
где и - параметры уравнения.
Для сообществ планктонных и донных животных были получены следующие значения параметров уравнений:
зоопланктон P b /R b = 0.888*e - 0.553 H , R 2 = 0.59 (32)
зообентос P b /R b = 0.771* e - 0.431 H , R 2 = 0.55 (33)
Такие количественные зависимости для двух важнейших подсистем экосистем водоемов дают возможность с достаточной степенью уверенности предположить, что и по отношению к экосистеме в целом следует ожидать такого же выражения количественной связи между структурными и функциональными характеристиками, т.е.:
(P/R) e = 1 - e - 1 H
Количественное выражение такой зависимости для экосистемы в целом может быть получено, поскольку показана возможность использования индекса Шеннона для оценки разнообразия, а значит и степени сложности экосистемы (Jizhong, Shijun, 1991). При этом следует принимать во внимание, что отношение биомассы зоопланктона и зообентоса возрастает по мере увеличения продуктивности водоемов (Алимов, 1990) и связано с соотношением первичной продукции планктона и макрофитов. По мере снижения доли макрофитов в первичной продукции водоема возрастает роль сообществ зоопланктона по отношению к сообществам бентоса (Винберг, Алимов и др., 1988). Это и понятно, так как озера с развитой подводной растительностью богаты детритом, который может активно потребляться животными бентоса. В большинстве водохранилищ развитие детритной трофической цепи обусловлено значительным поступлением в них аллохтонных органических веществ, что и обеспечивает хорошие условия для развития сообществ донных животных.
Использование индекса Шеннона для оценки сложности экосистемы в целом, рассчитываемого с учетом численностей конкретных видов вряд ли возможно, поскольку определение численности конкретных видов практически невозможно для сообществ бактерий и затруднено по отношению к водорослям планктона и особенно перифитона. Поэтому по отношению к экосистеме вероятно более надежно рассчитывать величину этого индекса с учетом биомассы отдельных групп гидробионтов:
H = (B i /B)*lg 2 (B i /B).
Для расчетов индексов разнообразия и отношения (P/R) e были использованы наиболее надежные и подробные биотические балансы, составленные для экосистем Нарочанских озер в 1972 и 1985 г.г., оз.Щучьего (1981, 1982 г.г.), оз.Красного, исландского оз. Thingvallavatn (Ecology of oligotrophic ...,1992). При этом были отобраны биотические балансы для этих озер только для тех лет наблюдений, когда Р е >0. Полученные результаты приведены на рис.26 и они могут быть аппроксимированы уравнением:
(P/R) e = 1.066*e - 2.048H , R 2 = 0.496 . (34)
Значения (Р/В) е - коэффициентов, как было показано (рис. 25) не зависят от продуктивности водоемов, но при этом наблюдается достаточно большой разброс данных. Отклонение значения коэффициента для экосистемы конкретного озера от средней для всех изученных экосистем (K = (P/B) e - (P/B) е среднее) связано, с достаточной степенью достоверности, со степенью сложности структуры экосистем (рис.27):
К = 0.902*Н - 0.778. R 2 = 0.561.
Таким образом, как и следовало, ожидать, структурные и функциональные характеристики сообществ гидробионтов и экосистем связаны между собой и эта связь может быть представлена в виде уравнений экспоненциальной функции: по мере усложнения структуры сообществ гидробионтов и экосистем возрастает доля рассеиваемой энергии в виде тепловой по отношению к энергии, заключенной в продукции этих биологических систем.
Из сказанного следуют два важных вывода. Во-первых, строгие количественные связи между структурными и функциональными характеристиками биосистем дают основание надеяться на получение количественных соотношений между потоками энергии и информации в водных экосистемах. Во-вторых, структура сообществ организмов и экосистем сохраняется не за счет установления устойчивых связей между элементами (как у объектов неживой природы), а за счет постоянных затрат энергии на поддержание упорядоченности и воспроизводства элементов системы, их структур и структур организмов.
Состояние сообществ гидробионтов и экосистем может быть описано с помощью структурных и функциональных характеристик. Изменение структуры системы связанное, например, с исчезновением отдельных видов, изменением трофических связей вызывает изменение функциональных характеристик системы и она переходит в новое состояние, определяемое новыми структурными и функциональными характеристиками.
Получение высокой продукции сообщества животных или экосистем возможно лишь при упрощении их структуры, в том числе в результате эксплуатации экосистем. Существенно, что продукция популяций определяется не только их продукционным потенциалом, величинами доступных животным кормовых ресурсов, но и интенсивностью эксплуатации популяции (Алимов, Умнов, 1989) или организацией определенной возрастной структуры популяции (Умнов, 1997).
Разная степень эксплуатации озерных экосистем приводит к изменениям их структурных и функциональных характеристик. Так, например, по мере увеличения нагрузки рыбами экосистем озер-рыбопитомников доля продукции сообществ донных животных в общих тратах энергии сначала увеличивается и, достигнув некоторого максимума, начинает снижаться (рис.28,). Это дает основание определить некоторый оптимальный режим эксплуатации таких озер. В обычных озерах, в которых ведется нормальный промысел рыб, как видно из рис. 26, доля продукции кормовых объектов закономерно снижается при возрастании пресса рыб. Причем такая закономерность наблюдается не только по отношению к сообществам донных животных, но и сообществ зоопланктона и бентоса. Отношение продукции и трат на обменные процессы в сообществах планктонных и бентосных животных, как кормовых объектов рыб, снижается по мере возрастания средней массы рыб в водоеме. Это означает, что в водоемах, где более крупные рыбы, в сообществах кормовых организмов доля энергии в продукции по отношению к рассеиваемой в процессах обмена энергии ниже, чем в водоемах с преобладанием рыб меньшего размера. Если вспомнить, что чем более сложно организовано сообщество животных, тем большая доля энергии рассеивается в процессах обмена по отношению к энергии, запасенной в продукции, можно предполагать, что увеличение среднего размера рыб в водоеме приводит к усложнению структуры сообществ их кормовых объектов. Такое явление может быть связано с тем, что рыбы потребляют в первую очередь животных крупных размеров и, тем самым снижают их доминирование в сообществах животных. Это подтверждает положение о том, что рыбы способствуют поддержанию высокого видового разнообразия и стабилизации потоков энергии в сообществах животных, высказанное Пейном (Paine, 1966).
На состав и распределение фитопланктона по отдельным водоемам, на его изменение в пределах одного водоема влияет большой комплекс факторов. Первостепенное значение из физических факторов имеют световой режим, температура воды, а для глубоких водоемов - вертикальная устойчивость водных масс. Из химических факторов основное значение имеют соленость воды и содержание в ней питательных веществ, в первую очередь солей фосфора, азота, а для некоторых видов также железа и кремния. Рассмотрим некоторые из перечисленных факторов.
Влияние освещенности как экологического фактора наглядно проявляется в вертикальном и сезонном распределении фитопланктона. В морях и озерах фитопланктон существует лишь в верхнем слое воды. Нижняя граница его в морских, более прозрачных водах находится на глубине 40-70 м и лишь в немногих местах достигает 100--120 м (Средиземное море, тропические воды Мирового океана). В озерных значительно менее прозрачных водах фитопланктон существует обычно в верхних слоях, на глубине 10-15 м, а в водах с очень малой прозрачностью встречается на глубине до 2-3 м. Лишь в высокогорных и некоторых крупных озерах (например, Байкале) с прозрачной водой фитопланктон распространен до глубины 20-30 м. Прозрачность воды в дант ном случае влияет на водоросли не прямо, а косвенно, поскольку она определяет интенсивность проникновения в водную толщу солнечной радиации, без которой невозможен фотосинтез. Это хорошо подтверждает сезонный ход развития фитопланктона в водоемах умеренных и высоких широт, замерзающих в зимний период. Зимой, когда водоем покрыт льдом, часто еще со слоем снега, несмотря на самую высокую в году прозрачность воды, фитопланктон почти отсутствует - встречаются лишь весьма редкие физиологически неактивные клетки некоторых видов, а у отдельных водорослей -- споры или клетки в стадии покоя.
При общей большой зависимости фитопланктона от освещенности оптимальные значения последней у отдельных видов варьируют в довольно широких пределах. Особенно требовательны к этому фактору зеленые водоросли и большинство видов сине-зеленых, в значительном количестве развивающихся в летний сезон. Некоторые виды сине-зеленых в массе развиваются только у самой поверхности воды: осциллатория (Oscillatoria) -в тропических морях, многие виды микроцистиса (Microcystis), анабены(Anabaena) и др. - в мелких внутренних водоемах.
Менее требовательны к условиям освещенности - диатомовые. Большинство из них избегает ярко освещенного приповерхностного слоя воды и более интенсивно развивается лишь на глубине 2-3 м в малопрозрачных водах озер и на глубине 10-15 м в прозрачных водах морей.
Температура воды - важнейший фактор общего географического распределения фитопланктона и сезонных циклов его, но действует этот фактор во многих случаях не прямо, а косвенно. Многие водоросли способны переносить большой диапазон колебаний температуры (эвритермные виды) и встречаются в планктоне разных географических широт и в разные сезоны года. Однако зона температурного оптимума, в пределах которого наблюдается наибольшая продуктивность, для каждого вида обычно ограничена не большими отклонениями температуры. Например, широко распространенная в озерном планктоне умеренной зоны и субарктики диатомея мелозира исландская (Melosira islandica) обычно присутствует в планктоне (например, в Онежском и Ладожском озерах, в Неве) при температуре от +1 до + 13°С, максимальное же размножение ее наблюдается при температуре от +6 до +8 °С.
Температурный оптимум у разных видов не совпадает, чем и определяется смена видового состава по сезонам, так называемая сезонная сукцессия видов. Общая, схема годового цикла фитопланктона в озерах умеренных широт имеет следующий вид. Зимой подо льдом (особенно когда лед покрыт снегом) фитопланктон почти отсутствует в связи с недостатком солнечной радиации. Вегетационный цикл фитопланктона как сообщества начинается в марте - апреле, когда солнечной радиации достаточно для фотосинтеза водорослей даже подо льдом. В это время бывают довольно многочисленными мелкие жгутиковые - криптомонас (Cryptomonas), хромулина (Chromulina), хризококкус (Chrysococcus) -- и начинается повышение численности холодноводных видов диатомовых -- мелозиры (Melosira), диатомы(Diatoma) и др.
Во вторую фазу весны -- с момента вскрытия льда на озере до установления температурной стратификации, что обычно бывает при прогреве верхнего слоя воды до +10, +12 °С, наблюдается бурное развитие холодноводного комплекса диатомовых. В первую фазу летнего сезона, при температуре воды от+10 до + 15 °С, холодноводный комплекс диатомовых прекращает вегетацию, в планктоне в это время еще многочисленны диатомовые, но уже другие виды - умеренно тепловодные: астерионелла (Asterionella), табеллария (Tabellaria). Одновременно повышается продуктивность зеленых и сине-зеленых водорослей, а также хризомонад, часть видов которых достигает значительного развития уже во вторую фазу весны. Во вторую фазу лета, при температуре воды выше + 15 °С, наблюдается максимум продуктивности сине-зеленых и зеленых водорослей. В зависимости от трофического и лимнологического типа водоема в это время может наблюдаться «цветение» воды, вызванное видами сине-зеленых (Anabaena, Aphanizomenon, Microcystis, Gloeotrichia, Oscillatoria) и зеленых водорослей (Scenedesmus, Pediastrum, Oocystis).
Диатомовые летом, как правило, занимают подчиненное положение и представлены тепловодными видами: фрагиларией (Fragilaria) и мелозирой (Melosira granulata). Осенью, с понижением температуры воды до +10, +12 °С и ниже, снова наблюдается подъем продуктивности холодноводных видов диатомовых. Однако, в отличие от весеннего сезона, в это время заметно большую роль играют сине-зеленые водоросли.
В морских водах умеренных широт весенняя фаза в фитопланктоне также выделяется вспышкой диатомовых водорослей; летняя же -- повышением видового разнообразия и обилия перидинеи при депрессии продуктивности фитопланктона в целом.
Из химических факторов, влияющих на распределение фитопланктона, на первое место следует поставить солевой состав воды. При этом общая концентрация солей является важным фактором качественного (видового) распределения по типам водоемов, а концентрация питательных солей, прежде всего солей азота и фосфора,- количественного распределения, т. е. продуктивности.
Общая концентрация солей нормальных (в экологическом смысле) природных вод варьирует в очень широких пределах: примерно от 5-10 до 36 000-38 000 мг/л (от 0,005-0,01 до 36--38°/0О). В этом диапазоне солености выделяются два основных класса водоемов: морские с соленостью 36--38°/00, т. е.36 000-- 38 000 мг/л, и пресные с соленостью от 5--10 до 400--500 и даже до 1000 мг/л. Промежуточное положение по концентрации солей занимают солоноватые воды. Этим классам вод, как было показано выше, соответствуют и основные группы фитопланктона по видовому составу.
Экологическое значение концентрации биогенных веществ проявляется в количественном распределении фитопланктона в целом и составляющих его видов.
Продуктивность, или «урожайность», микроскопических водорослей фитопланктона, как и урожайность крупной растительности, при прочих нормальных условиях в очень большой степени зависит от концентрации питательных веществ в окружающей среде. Из минеральных питательных веществ для водорослей, как и для наземной растительности, в первую очередь необходимы соли азота и фосфора. Средняя концентрация этих веществ в большинстве естественных водоемов очень мала, и поэтому высокая продуктивность фитопланктона, как устойчивое явление, возможна лишь при условии постоянного поступления минеральных веществ в верхний слой воды -- в зону фотосинтеза.
Правда, некоторые сине-зеленые водоросли способны еще усваивать элементарный азот из растворенного в воде воздуха, однако таких видов немного и их роль в обогащении азотом бывает существенной лишь для очень мелких водоемов, в частности на рисовых полях.
Внутренние водоемы удобряются азотом и фосфором с берега, за счет приноса питательных веществ речной водой с водосборной площади всей речной системы. Поэтому наблюдается четкая зависимость продуктивности озер и мелководных внутренних морей от плодородия почв и некоторых других факторов, действующих в пределах водосборной площади их бассейнов (речных систем). Наименее продуктивен фитопланктон при ледниковых озер, а также водоемов, расположенных на кристаллических породах и в районах с большим количеством болот в пределах водосборной площади. Примером последних могут служить озера Северной Карелии, Кольского полуострова, Северной Финляндии, Швеции и Норвегии. Наоборот, водоемы, расположенные в пределах высокоплодородных почв, отличаются высоким уровнем продуктивности фитопланктона и других сообществ (Азовское море, нижневолжские водохранилища, Цимлянское водохранилище).
Продуктивность фитопланктона зависит и от динамики воды, динамического режима вод. Влияние может быть прямым и косвенным, что, однако, не всегда легко различить. Турбулентное перемешивание, если оно не слишком интенсивно, при прочих благоприятных условиях прямо способствует повышению продуктивности диатомовых водорослей, так как многие виды этого отдела, обладая относительно тяжелой оболочкой из кремния, в спокойной воде опускаются на дно. Поэтому ряд массовых пресноводных видов, в частности из рода мелозира, интенсивно развиваются в планктоне озер умеренных широт лишь весной и осенью, в периоды активного вертикального перемешивания воды. При прекращении такого перемешивания, наступающем при прогреве верхнего слоя до +10, +12 °С и образовании при этом во многих озерах температурного расслоения водной толщи, эти виды из планктона выпадают.
Другие водоросли, прежде всего сине-зеленые, наоборот, не выносят даже относительно слабого турбулентного перемешивания воды. В противоположность диатомовым многие виды сине-зеленых наиболее интенсивно развиваются в предельно спокойной воде. Причины высокой чувствительности их к динамике вод не вполне установлены.
Однако в тех случаях, когда вертикальное перемешивание вод распространяется на большую глубину, оно подавляет развитие даже относительно теневыносливых диатомовых. Связано это с тем, что при глубоком перемешивании водоросли периодически выносятся токами воды за пределы освещенной зоны - зоны фотосинтеза.
Косвенное влияние динамического фактора на продуктивность фитопланктона состоит в том, что при вертикальном перемешивании воды питательные вещества поднимаются из придонных слоев воды, где они не могут быть использованы водорослями вследствие недостатка света. Здесь проявляется взаимодействие нескольких экологических факторов - светового и динамического режимов и обеспеченности питательными веществами. Такая взаимосвязь характерна для природных процессов.
Уже в начале нашего века гидробиологи открыли особое значение фитопланктона в жизни водоемов как основного, а на обширных океанических просторах и единственного производителя первичного органического вещества, на базе которого создается все остальное многообразие водной жизни. Это определило повышенный интерес к изучению не только качественного состава фитопланктона, но и количественного распределения его, а также факторов, регулирующих это распределение.
Элементарный метод количественной оценки фитопланктона, который на протяжении нескольких десятилетий был основным, да и теперь еще не полностью отвергнут, - метод отцеживания его из воды с помощью планктонных сеток. В сконцентрированной таким путем пробе просчитывают количество клеток и колоний по видам и определяют общую численность их на единицу поверхности водоема. Этот простой и доступный метод имеет, однако, существенный недостаток -- он не полностью учитывает даже относительно крупные водоросли, а самые мелкие (наннопланктон), которые во многих водоемах значительно преобладают, планктонные сетки не улавливают.
В настоящее время пробы фитопланктона берут в основном батометром или планктобатометром, позволяющим «вырезать» монолит воды с заданной глубины. Сгущение пробы производится методом осаждения в цилиндрах или фильтрацией через микрофильтры: то и другое гарантирует учет водорослей всех размеров.
Когда определились огромные различия в размерах водорослей, составляющих фитопланктон (от нескольких до 1000 мкм и более), стало ясно, что для сравнительной оценки продуктивности фитопланктона по водоемам величинами численности пользоваться нельзя. Более реальным показателем для этой цели является общая биомасса фитопланктона на единицу площади водоема. Однако в дальнейшем и этот метод был забракован по двум основным причинам: во-первых, расчеты биомассы клеток, имеющих у разных видов разную конфигурацию, очень трудоемки; во-вторых, вклад мелких, но быстро размножающихся водорослей в общую продукцию сообщества за единицу времени может быть значительно большим, чем крупных, но медленно размножающихся.
Истинным показателем продуктивности фитопланктона является скорость образования им вещества за единицу времени. Для определения этой величины пользуются физиологическим методом. В процессе фотосинтеза, происходящем только на свету, поглощается углекислота и выделяется кислород. Наряду с фотосинтезом происходит и дыхание водорослей. Последний процесс, связанный с поглощением кислорода и выделением углекислоты, превалирует в темноте, когда фотосинтез прекращается. Метод оценки продуктивности фитопланктона основан на количественном сопоставлении результатов фотосинтеза (процесса продукции) и дыхания (процесса деструкции) сообщества по балансу кислорода в водоеме. Для этой цели используются пробы воды в светлых и темных склянках, экспонируемых в водоеме обычно на сутки на разных глубинах.
Для повышения чувствительности кислородного метода, непригодного для малопродуктивных вод, стали применять изотопную (радиоуглеродную) разновидность его. Однако впоследствии выявились недостатки кислородного метода в целом, и в настоящее время широко применяют хлорофилльный метод, основанный на определении содержания хлорофилла в количественной пробе фитопланктона.
В настоящее время уровень продуктивности фитопланктона многих внутренних водоемов определяется не столько природными условиями, сколько общественно-экономическими, т. е. плотностью населения и характером хозяйственной деятельности в пределах водосборной площади водоема. Эта категория факторов, именуемая в экологии антропогенными, т. е. происходящими от деятельности человека, приводит к обеднению фитопланктона в одних водоемах, а в других, наоборот, к значительному повышению его продуктивности. Первое происходит в результате сброса в водоем токсических веществ, содержащихся в сточных водах промышленного производства, а второе - при обогащении водоема биогенными веществами (особенно соединениями фосфора) в минеральной или органической форме, содержащимися в больших концентрациях в водах, стекающих с сельскохозяйственных территорий, из городов и мелких селений (бытовые стоки). Биогены содержатся и в сточных водах многих промышленных производств.
Второй вид антропогенного влияния - обогащение водоема биогенными веществами - повышает продуктивность не только фитопланктона, но и других водных сообществ, до рыб включительно, и его следовало бы рассматривать как благоприятный с экономической точки зрения процесс. Однако во многих случаях стихийное антропогенное обогащение водоемов первичными питательными веществами происходит в таких масштабах, что водоем как экологическая система оказывается перегруженным биогенами. Следствием этого является чрезмерно бурное развитие фитопланктона («цветение» воды), при разложении которого выделяется сероводород или другие токсические вещества. Это приводит к гибели животного населения водоема и делает воду непригодной для питья.
Нередки случаи и прижизненного выделения водорослями токсических веществ. В пресноводных водоемах чаще всего это наблюдается при массовом развитии сине-зеленых водорослей, в частности видов рода микроцистис (Microcystis). В морских водах отравление воды нередко вызывается массовым развитием мелких жгутиковых. В таких случаях вода иногда окрашивается в красный цвет, отсюда и название этого явления - «красный прилив».
Понижение качества воды в результате антропогенной перегрузки водоема биогенными веществами, вызывающей чрезмерное развитие фитопланктона, принято называть явлением антропогенной эвтрофикации водоема. Это одно из печальных проявлений загрязнения окружающей среды человеком. О масштабах этого процесса можно судить по тому, что загрязнение интенсивно развивается в таких огромных пресных водоемах, как озеро Эри, и даже в некоторых морях.
Естественное плодородие морских поверхностных вод определяется разными факторами. Пополнение питательными веществами мелководных внутренних морей, например Балтийского, Азовского, происходит в основном за счет приноса их речными водами.
Поверхностные воды океанов обогащаются питательными веществами в районах выхода глубинных вод на поверхность. Явление это вошло в литературу под названием апвеллинга. Очень интенсивен апвеллинг у перуанского побережья. На базе высокой продукции фитопланктона здесь чрезвычайно высока продукция беспозвоночных, а за счет этого растет численность рыб. Небольшая страна, Перу в 60-х годах по уловам рыбы вышла на первое место в мире.
Мощная продуктивность фитопланктона в холодных водах арктических морей и особенно в водах Антарктики определяется также подъемом глубинных вод, обогащенных биогенными веществами. Подобное явление наблюдается и в некоторых других районах океана. Противоположное явление, т. е. обеднение поверхностных вод питательными веществами, тормозящее развитие фитопланктона, наблюдается в районах с устойчивой изоляцией поверхностных вод от глубинных.
Таковы основные особенности типичного фитопланктона.
Растения наших водоемов: фитопланктон
Фитопланктоном называют микроскопические водоросли, свободно «парящие» в толще воды. Для жизни в таком состоянии в процессе эволюции у них выработался ряд приспособлений, которые способствуют уменьшению относительной плотности клеток (накопление включений, образование газовых пузырьков) и увеличению их трения (отростки различной формы, выросты).
Пресноводный фитопланктон представлен в основном зелеными, сине-зелеными, диатомовыми, пирофитовыми, золотистыми и эвгленовыми водорослями.
Развитие фитопланктонных сообществ происходит с определенной периодичностью и зависит от различных факторов. Например, прирост биомассы микроводорослей до определенного момента происходит пропорционально количеству поглощаемого света. Зеленые и сине-зеленые водоросли наиболее интенсивно размножаются при круглосуточном освещении, диатомовые - при более коротком фотопериоде. Начало вегетации фитопланктона в марте-апреле в немалой степени связано с повышением температуры воды. Диатомовым свойственен низкий температурный оптимум, для зеленых и сине-зеленых - более высокий. Поэтому весной и осенью при температуре воды от 4 до 15 градусов в водоемах доминируют диатомовые водоросли. Увеличение мутности воды, вызываемое минеральными взвесями, снижает интенсивность развития фитопланктона, особенно сине-зеленых. Менее чувствительны к повышению мутности воды диатомовые и протококковые водоросли. В воде, богатой нитратами, фосфатами и силикатами, развиваются преимущественно диатомовые, в то же время зеленые и сине-зеленые менее требовательны к содержанию этих биогенных элементов.
На видовой состав и численность фитопланктона оказывают влияние и продукты жизнедеятельности самих водорослей, поэтому между некоторыми из них существуют, как отмечается в научной литературе, антагонистические взаимоотношения.
Из всего многообразия видов пресноводного фитопланктона диатомовые, зеленые и сине-зеленые водоросли - наиболее многочисленны и особенно ценны в кормовом отношении.
Клетки диатомовых водорослей снабжены двустворчатой оболочкой из кремнезема. Их скопления отличаются характерной, желтовато бурой окраской. Эти микрофиты играют важную роль в питании зоопланктона, но из-за низкого содержания органического вещества их пищевая ценность не столь значительна как, например, у протококковых водорослей.
Отличительный признак зеленых водорослей - типичная зеленая окраска. Их клетки, содержащие ядро и хроматофор, различны по форме, часто снабжены шипами и щетинками. Некоторые имеют красный глазок (стигма). Из представителей этого отдела протококковые водоросли являются объектами массового культивирования (хлорелла, сценедесмус, анкистродесмус). Их клетки отличаются микроскопическими размерами и легко доступны фильтрующим гидробионтам. Калорийность сухого вещества этих водорослей приближается к 7 ккал/г. В них много жира, углеводов, витаминов.
Клетки сине-зеленых водорослей не имеют хроматофоров и ядер и равномерно окрашены в сине-зеленый цвет. Иногда их окраска может приобретать фиолетовый, розовый и другие оттенки. Калорийность сухого вещества достигает 5,4 ккал/г. Белок полноценен по аминокислотному составу, однако из-за слабой растворимости он малодоступен для рыб.
В создании естественной кормовой базы водоемов фитопланктону принадлежит ключевая роль. Микрофиты как первичные продуценты, усваивая неорганические соединения, синтезируют органические вещества, которые утилизируются зоопланктоном (первичный консумент) и рыбами (вторичный консумент). От соотношения крупных и мелких форм в фитопланктоне в значительной мере зависит и структура зоопланктона.
Один из факторов, лимитирующих развитие микрофитов,- содержание в воде растворимого азота (преимущественно аммонийного) и фосфора. Для прудов оптимальной нормой считают 2 мг N/л и 0,5 мг Р/л. Увеличению биомассы фитопланктона способствует дробное внесение за сезон 1 ц/га азотно-фосфорных, а также органических удобрений.
Продукционные возможности водорослей достаточно велики. Применяя соответствующую технологию, с 1 га водной поверхности можно получать до 100 т сухого вещества хлореллы.
Промышленное культивирование водорослей слагается из ряда последовательных этапов с использованием различного рода реакторов (культиваторов) на жидких средах. Средняя урожайность водорослей, по данным ВНИИПРХа, колеблятся от 2 до 18,5 г сухого вещества на 1 м: в сутки.
Мерой продуктивности фитопланктона служит скорость образования органического вещества в процессе фотосинтеза. Врдоросли - основной источник первичной продукции. Первичная продукция - количество органического вещества, синтезируемого эвтрофными организмами за единицу времени,- обычно выражается в ккал/ м в сутки.
Фитоплактон наиболее точно определяет трофический уровень водоема. К примеру, для олиготрофных и мезотрофных вод характерно низкое отношение численности фитопланктона к его биомассе, а для гипертрофных - высокое. Биомасса фитопланктона в гипертрофных водоемах составляет более 400 мг/л, в эвтрофных - 40,1-400 мг/л, в дистрофных - 0,5-1 мг/л.
Антропогенная эвтрофикация - возросшее насыщение водоема биогенами - одна из злободневных проблем. Определить степень активности биологических процессов в водоеме, как и степень его интоксикации, можно с помощью фитопланктонных организмов - индикаторов сапробности. Различают водоемы поли-, мезо- и олигосапробные. По классической системе организмов-индикаторов, созданной Кольквитцем и Марссоном, к полисапробионтам можно, например, отнести Euglena viridis, к альфа-мезосапробионтам - Chlorella vulgaris, к олигосапробионтам - Sy-nura uvella. Поскольку методы гидробиологического контроля качества воды постоянно совершенствуются, список организмов-индикаторов сапробности расширяется и уточняется.
Повышение эвтрофикации, или чрезмерное накопление в водоеме органического вещества, тесно связано с усилением процессов фотосинтеза в фитопланктоне. Массовое развитие водорослей приводит к ухудшению качества воды, ее «цветению».
Цветение - не стихийное явление, оно подготавливается в течение довольно продолжительного времени, иногда двух и более вегетационных периодов. Предпосылки резкого возрастания численности фитопланктона - наличие водорослей в водоеме и их способность к размножению при благоприятных условиях. Развитие диатомовых, например, в значительной мере зависит от содержания в воде железа, лимитирующим фактором для зеленых водорослей служит азот, сине-зеленых - марганец. Цветение воды считается слабым, если биомасса фитопланктона находится в пределах 0,5-0,9 мг/л, умеренным - 1-9,9 мг/л, интенсивным - 10- 99,9 мг/л, а при гиперцветении она превышает 100 мг/л.
Методы борьбы с этим явлением пока еще не настолько совершенны, чтобы можно было считать проблему окончательно решенной. Из гидрологических мер наиболее часто употребляемые и безопасные - увеличение проточности и искусственная аэрация водоема.
В качестве альгицидов (химических средств борьбы с цветением) применяют производные карбамида - диурон и монурон - в дозах 0,1-2 мг/л. Для временной очистки отдельных участков водоемов вносят сернокислый алюминий. Однако прибегать к ядохимикатам следует с осторожностью, так как они потенциально опасны не только для гидробионтов, но и для человека.
В последние годы в этих целях широко используют растительноядных рыб. Так, белый толстолобик потребляет различные виды протококковых, эвгленовых, диатомовых водорослей. Сине-зеленые, продуцирующие при массовом развитии токсические метаболиты, усваиваются им хуже, однако в рационе взрослых особей этой рыбы они могут составлять значительную долю. Фитопланктон охотно поедают также тиляпия, серебряный карась, пестрый толстолобик, а при недостатке основной пищи - сиговые, большеротый буффало, веслонос.
В определенной мере ограничивать интенсивность цветения воды могут и макрофиты. Помимо выделения в воду вредных для фитопланктона веществ, они затеняют поверхность близлежащих участков, препятствуя фотосинтезу.
При расчете кормовой базы водоема и продукции фитопланктона приходится определять видовой состав, численность клеток и биомассу водорослей по содержанию в определенном объеме воды (0,5 или 1 л).
Методика обработки пробы включает в себя несколько этапов (фиксация, концентрирование, приведение к заданному объему). Существует много различных фиксаторов, однако чаще всего употребляется формалин (2-4 мл 40 %-ного раствора формалина на 100 мл воды). Клетки водорослей отстаивают в течение двух недель (если объем пробы меньше 1 л, соответственно укорачивается и период осаждения). Затем верхний слой отстоявшейся воды осторожно удаляют, оставляя для дальнейшей работы 30-80 мл.
Клетки фитопланктона подсчитывают небольшими по объему порциями (0,05 или 0,1 мл), затем по полученным результатам определяют их содержание в 1 л. Если численность клеток того или иного вида водорослей превышает 40 % от их общего количества, то данный вид считается доминирующим.
Определение биомассы фитопланктона - трудоемкий и длительный процесс. На практике для облегчения расчета условно принято считать, что масса 1 млн. клеток пресноводного фитопланктона приблизительно равна 1 мг. Есть и другие экспресс-методы. Учитывая большую роль фитопланктона в экосистеме водоемов, в формировании их рыбопродуктивности, необходимо, чтобы этими методами владели все рыбоводы - от ученых до практиков.
С. Юдин
Продукционный цикл фитопланктона
Основными факторами, определяющими первичную продукцию фитопланктона, являются толщина фотического слоя, обеспеченность фитопланктона биогенными элементами и продолжительность продукционного цикла.
В целом указанные параметры зависят от конкретных климатических и гидрологических условий каждого района и проявляются в сроках наступления весеннего развития, которые тесно связаны следовым режимом и окончания продукционного цикла; кроме того, эти параметры определяются запасом биогенных элементов, образовавшимся в фотическом слоек началу весеннего развития и определяемым речным стоком и глубиной зимнего конвективного перемешивания, летней вертикальной турбулентной диффузией, количеством солнечной энергии (зависящей в свою очередь от времени года и климатических факторов), а также структурно-функциональными характеристиками фитопланктона.
Фитопланктон Белого моря представляет собой качественно обедненный планктон Баренцева моря с большой примесью «литоральных» эпифитных и пресноводных форм (Кокин, Кольцова, 1972). В Белом море по сравнению с южной частью Баренцева моря по числу видов преобладают холодолюбивые диатомеи и снижается видовое разнообразие перидиней. Подоле содержания водорослей в составе фитопланктона (63%) Белое море приближается к таким высокоарктическим эпиконтинентальным морям, как Карское (67%) и Лаптевых (64 %), что лишний раз указывает на его арктический характер. Формирование качественного и количественного состава фитопланктона в различные биологические сезоны происходит под влиянием как биотических, так и абиотических факторов среды.
Изменение структурных и функциональных показателей сообщества в течение продукционного цикла можно проследить на примере анализа биологических сезонов в Кандалакшском заливе в 1970 г. (Федоров, Бобров, 1977, Бобров, 1982).
Всего за период наблюдений с 15 апреля по 26 октября в 1970 г. поставлена 51 станция (дважды в неделю). Продукцию определили радиоуглеродным методом склянок на горизонтах 0,2.5, 5, 10, 25, 50, 75 м и у дна. Суммарную солнечную радиацию измеряли пиранометром Янышевского в подводной модификации. Одновременно с постановкой опытов по определению первичной продукции с соответствующих горизонтов проводили батометрические количественные сборы фитопланктона и изучение некоторых гидролого-гидрохимических параметров. При расчете эффективности фотосинтетического процесса данные измерения первичной продукции были выражены в энергетических единицах, исходя из следующего расчета: на фиксацию I моля CO2 расходуется 112 калорий световой энергии, а эта величина соответствует 9.36 кал/мгC.
Переход от численности особей фитопланктона к весовым характеристикам производили по формуле:
Формула получена с учетом того, что в сухой биомассе водорослей содержится 35 % золы (Федоров и др., 1974).
Величины суточных сумм радиации в течение вегетационного периода 1970 г. колебались от 20.6 до 2300 кал/(см". день). Увеличение проникающей в BOAy солнечной радиации в первой декаде мая до 100.6-181.4 кал/(см" - день) совпадает повремени с вскрытиемледового покрова. Некоторое снижение значений суммарной солнечной энергии было вызванометеорологическими условиями Улучшение погодных условий и увеличение длины светового дня привело к практическому совпадению светового максимума (2300 кал/(см". день)) с календарным днем летнего солнцестояния. Последующее падение дневных сумм солнечной радиации имеет довольно плавный характер и обусловливается уменьLEHI HEM продолжительности CBPTOBOTO IH H H BECOTE CONHL kl. К концу вегетационного сезона величина солнечной энергии падает до 20 кал/(см". день).
Таким образом, за весь период наблюдений, охватывающий 183 дня, величина энергии выразилась суммарным значением 22.8 ккал/см". Неучтенная энергия проникающей солнечной радиации в период осении «подледной весны» была получена экстраполяцией и составила 22 ккал/см". Всего за сезон в водоем проникло 25 ккал/см". Последнее значение можно считать годовой суммой, поскольку в зимние месяцы количество энергии солнечной радиации, проникающей под ледяной покров, незначительно, Изучение распространения солнечной энергии в толще воды на протяжении вегетационного периода позволило рассчитать коэффициент поглощения света водой k=0.37-002.
Температура в поверхностном горизонте в течение вегетационного сезона колебалась от - 1.54 до 15.65 °С. С глубиной температурные колебания сглаживались. Весенний период после вскрытия льда характеризуется нарушением гомотермии и установлением отчетливо выраженной стратификации. Зона температурного скачка с ростом проникающей солнечной энергии и соответствующим прогревом ВМ постепенно перемещалась из поверхностного в более глубокие слои. В период максимального значения солнечной энергии термоклин располагался на глубине 25 м и удерживался на этом горизонте до конца сентября. В октябре температурный скачок был расположен глубже и обнаруживалась тенденция к гометермии.
В табл. 5.1 представлены некоторые абиотические характеристики Белого моря, полученные на основе анализа их сезонной LHHaMHK.H.
Анализ вегетационного развития планктона и его продукционHEIM Xapa KTepH cTH K позволяет выделить и установить продолжительность основных периодов вегетации, названных В. Г. Богоровым (1938) биологическими весной, летом и осенью. Для Белого моря характерны все три периода. Их продолжительность для Кандалакшского залива в 1970 г. составила 48, 92 и 43 сут соответственно.
Результаты, характеризующие абсолютные величины продукционного процесса в различные периоды вегетации, представлены в табл. 5.2. Коэффициент Р/Сент рассчитывали как отношение первичной продукции к биомассе (выраженной в углероде) для Bice TO CTO,JI (5a B OIE bil.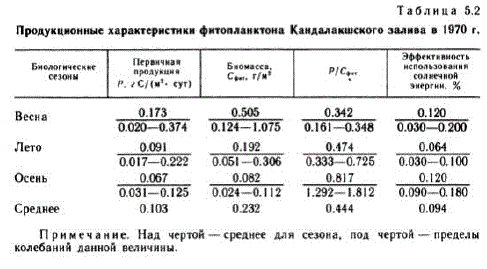
Примечание. Над чертой - среднее для сезона, под чертой - пределы колебаний данной величины.
Начало биологической весны отмечено впервой декаде апреля, Весенний подледный климакс (Федоров, 1970a; Кокин, Кольцова, 1972) продолжается около 2 недель и характеризуется устойчивым P/Cфи-коэффициентом. Распределение экониш водорослей соответствует второй модели Мак-Артура (Кокин, Кольцова, 1972).
Весенняя вспышка величин биомассы и численности водорослей начинается подо льдом и после вскрытия льда достигает максимума. Фитопланктон представлен BH La MH JULHETOMOBOTO KOM TIJEKCE: Fragilaria oceanica, Chaetoceros holsa ficus, Nitzschia frigida, Maticula septentrionaliа. Самым массовым видом была F. oceапіса. Фотический слой обогащен биогенными элементами и содержит 86 % всей биомассы фитопланктона, которая, так же как и скорость фотосинтеза, в этот период достигает максимального значения за весь вегетационный период (см. табл. 5.2). Выедание водорослей зоопланктоном (Конопля, 1973) приводит к падению биомассы водорослей и снижению пищевой конкуренции за биогенные элементы, при этом распределение водорослей соответствует уже первой модели Мак-Артура.
Летний максимум развития фитопланктона (первый), обусловленный увеличением числа диатомовых, после спада их численности «подхватывается» перидиниевыми водорослями, образующими второй летний максимум. С середины июля и до конца августа устанавливается летний климакс с устойчивым P/Cфиткоэффициентом. Доминирующими формами биологического лета оказываются Sceletoпeтa costalит и виды рода Chaetoceros (главным образом Ch. сотргеssиs). Первичная продукция в этот период характеризуется средней величиной 0,091 г С/(м", сут), а биомасса - 0.192 гC/м". Развита трофическая конкуренция за биогенные элементы,
Осенняя вспышка фитопланктона выражена слабо. Она наблюдается сконца августа до середины сентября и поросту биомассы ee, Rb Ae/y!"b, "pyAhQ. Только увеличение первичной продукции до 0.125 г С/(м", сут) и Р/Сеит-коэффициента (до 1812) в 20-х числах сентября указывает на кратковременную активацию фитопланктона. В этот период наблюдается развитие Dinophyes пorwegica - холодолюбивого доминанта биологической осени (Федоров, 1969). Период осенней сукцессии приводит экосистему к кратковременному климаксу, который длится вплоть до конца сентября. Дальнейшее снижение температуры и солнечной радиации, а также увеличение слоя перемешивания в связи с нарушением скачка плотности приводит к перераспределению биомассы фитопланктона по глубине и преобладанию процессов деструкции над продукцией.
В целом такой ход продукционного цикла фитопланктона можно ожидать и в других районах моря. Возможные различия будут выражены в сроках начала и конца вегетации фитопланктона, а также в некоторых различиях качественного состава доминирующих видов водорослей (Житина, 1981), что несомненно отразится на результатах продукционных процессов фитопланктона этих частей акватории.
Данные исследований продукционного цикла фитопланктона в 1970 г. позволили уточнить классификацию трофности Белого моря. В качестве дополнительной характеристики была использована эффективность использования солнечной энергии фитопланктоном (см. табл. 5.2), являющаяся интегральной характеристикой состояния фитопланктона, зависящая от комплекса условий окружающей среды и не обнаруживающая прямой связи с биомассой фитопланктона (Федоров, Бобров, 1977).
Если сравнить расчетные значения основных продукционных характеристик фитопланктона с известными данными для различных по продуктивности районов океана (Сорокин, 1973), то можно сделать вывод, что Кандалакшский залив относится к малопродуктивным районам с уровнем продукции фитопланктона 0,05- 0.10 г С/(м*- cут) и эффективностью использования солнечной энергии 0.04 %, характерной для олиготрофных вод. Следовательно, Кандалакшский залив, согласно классификации О. И. Кобленц-Мишке (Кобленц-Мишке и др., 1970), можно отнести к 1-му классу трофности с очень незначительным перемешиванием Глубинных вод.
В таблицах 6.7-6.9 суммированы результаты измерений первичной продукции в водах разных рифов. Как видно из таблиц, первичная продукция сильно варьирует даже в пределах вод одного и того же рифа. Однако эти последние колебания вполне закономерны и связаны прежде всего с изменениями концентрации фитопланктона на разрезах от мелководных зон рифа в сторону центра лагуны.
В лагунах атоллов с низкой первичной продукцией (атоллы о-ва Лау и Маршало-вых островов, островов Туамоту) нижний предел (2-4 мг С/м 3 в сутки) близок к ее величинам в омывающих их олиготрофных тропических водах пассатных течений, где она составляет обычно 1-5 мг С/м 3 в сутки (Сорокин, 1971а). Однако и в таких атоллах в водах их лагун возможны сезонные вспышки развития фитопланктона, которые могут достигать силы "цветения" (Sournia, Ricard, 1976а). Как видно из данных табл. 6.7, в лагунах атоллов, даже в тех из них, которые омываются олиготрофными тропическими водами с крайне низким уровнем развития фитопланктона, первичная продукция чаще всего соответствует среднему, а иногда и верхнему уровню продуктивности фитопланктона в мезотрофных морских бассейнах (15-50 мг С/м 3 в сутки). В лагуне крупного кольцевого рифа Каргадос-Карахос, расположенного в стрежне очень малопродуктивных вод Южного пассатного течения Индийского океана, первична продукция достигала 250 мг С/м 3 в сутки при биомассе водорослей (преимущественно диатомей) более 5,2 г/м 3 (табл. 6.10). Высокая продукция (70100 мг С/м 3 в сутки) была зарегистрирована в закрытой лагуне атолла Фаннинг (острова Лау) в период развития там динофлагеллят и кокковидных синезеленых водорослей (Gordon et al., 1971). Этот атолл также располагается в олиготрофных водах пассатного течения. Таким образом, даже в лагунах отдельных кольцевых рифов, расположенных в потоке олиготрофных вод, могут существовать достаточно высокопродуктивные сообщества фитопланктона, продукция и биомасса которых на 1-2 порядка выше, чем в омывающих их водах океана.
Судя по результатам имеющихся немногочисленных измерений, концентрации фотосинтетически активного хлоофилла "а" в воде лагун кольцевых рифов открытого океана в среднем в 5-10 раз выше, чем в окружающих водах океана. По данным единичных измерений, она варьировала в лагунах разных рифов в пределах от 0,2 до 1 мг/м 3 . В лагуне атолла Такапото, по результатам длительных наблюдений (Souгnia, Ricard, 1976а) , концентрация хлорфилла менялась в пределах от 0,26 до 0,77 мг/м 3 , причем она заметно варьировала и в разных частях лагуны в зависимости от интенсивности поступления в них вод океана во время приливов.
Существенный вклад в изучение фитопланктона и его продукции дали сезонные наблюдения, выполненные в водах барьерных и береговых рифов вблизи крупных островов, где были основаны биологические станции доя изучения жизни рифов и прилегающей неритической зоны океана. В числе таких хорошо изученных рифов следует упомянуть рифы у о-ва Носи Бе и у Тулеара (Мадагаскар), Гавайский риф у о-ва Оаху, Б. Барьерный риф Австралии, рифы Вапрао, Тиаура и Фааа на островах Таити и Муреа, рифы центрального Вьетнама вблизи г. Нячанг. Результаты измерения первичной продукции и биомассы фитопланктона барьерных и береговых рифов обобщены в табл. 6.8, 6.9, 6.10: Представленные в них данные очень сильно варьируют почти для всех изученных рифов. Они отражают закономерные измерения первичной продукции в разных зонах одного и того же рифа. При этом наименьшие величины ее (так же как и наименьшие величины биомассы фитопланктона: рис. 6.10) соответствуют мелководным зонам рифа, где вода находится в интенсивном контакте с донным населением и альгофлорой рифа. В этих зонах (флет, патч-рифы, литораль) фитопланктон, как правило, бывает обедненным и его продукция здесь резко падает. Максимальные величины продукции и биомассы фитопланктона, представленные в указанных таблицах, на каждом рифе соответствуют зонам лагуны или внешнего склона окаймляющего или берегового рифа с глубинами более 15-20 м, где вода находится вне прямого контакта с донным населением рифа. Величины первичной продукции в этих зонах, как правило, бывают значительно выше, чем в водах прилегающей акватории открытого моря. В среднем они выше, чем в воде лагун атоллов и кольцевых рифов открытого океана.
Высокие величины первичной продукции, достигающие уровня евтрофных вод, были отмечены в воде ряда береговых и барьерных рифов Индийского океана, таких, как рифы у Мадагаскара, у Сейшельских островов и у о-ва Сокотра, а также в лагуне Б. барьерного рифа в периодах "цветения" синезеленых водорослей (см. рис. 6.9). Суточная продукция достигала здесь 50-300 мг С/м 3 в сутки в верхнем слое воды, или 0,7-2,8 г С/м 3 . При этом в водах барьерных рифов островов, омываемых олиготрофными водами океана (Танги, Оаху, Лизард), первичная продукция в лагунах может достигать 50-60 мг С/м 2 , или 0,4-0,7 г углерода в 1 м 2 в сутки, что соответствует верхнему уровню продуктивности мезотрофных вод. В рифовых зонах побережья Центрального Вьетнама и его островов в Южно-Китайском море (острова Тху) и в Сиамском заливе (острова Намзу) (см. табл. 6.8, рис. 6.10) величины первичной продукции фитопланктона были высокими, несмотря на то что наблюдения производились в период сезонного минимума фитопланктона (Сорокин и др., 1982). Наибольшие величины первичной продукции, как правило, наблюдались в водах у внешнего края рифа на расстоянии 10-100 м от него над глубинами 5-15 м (см. рис. 6.10). Она обычно превышала здесь 30-40 мг С/м 3 в сутки. На отдельных разрезах в этой зоне в 1984 г. была зарегистрирована первичная продукция выше 100 мг С/м 3 в сутки.
Сведения о биомассе фитопланктона береговых и барьерных рифов пока еще весьма отрывочны. Наиболее подробные измерения биомассы фитопланктона в течение сезона были выполнены Анго (Angot, 1968) у о-ва Носи Бе (Мадагаскар). В.обобщенном виде результаты этих измерений приводятся на рис. 6.9. В табл. 6.10 представлены имеющиеся в литературе данные о биомассе фитопланктона ряда крупных рифов Индийского и Тихого океана. Как видно из таблицы, биомасса фитопланктона в водах барьерных и береговых рифов может достигать 3-8 г/м 3 . Самые низкие виличины ее были отмечены в воде береговых и барьерных рифов центральной части Тихого океана, находившихся в угнетенном состоянии (рифы у островов Панарора и Паама, Новые Гебридах, риф у о-ва Тувута, острова Лау).
Столь же ограниченны сведения о содержании хлорофилла "а" в водах барьерных и береговых рифов (табл. 6.11). Наиболее подробные ее измерения были выполнены в водах лагун островов Муреа и Таити французскими исследователям (Ricard, 1976; Sournia, Ricard, 1976). Эти измерения показали, что концентрация хлорофилла в них в разные сезоны года варьирует в пределах 0,03-0,17 мг/м 3 . В лагуне Гавайского рифа у остра Оаху, а также в лагуне Б. барьерного рифа зимой концентрация хлорофилла была близка к таковой в мезотрофных морских водах: 0,1-0,3 мг/м 3 . В периода осеннолетних максимумов развития фитопланктона в лагуне Б. барьерного рифа, а также в водах рифа у о-ва Носи Бе (Мадагаскар) концентрация хлорофилла в воде была выше на 1 мг/м 3 , что близко к его содержанию в евтрофных водах (Angot, 1968). Установлено влияние взмучивания всего слоя донных осадков, обогащенного микробентосом, в зоне волнобоя на концентрацию хлорофилла в водах рифа (Walker, 1981).
Как уже отмечалось выше, величины первичной продукции и биомассы фитопланктона сильно варьируют в разных зонах одного и того же рифа. В большинстве случаев самые низкие величины биомассы и продукции фитопланктона характерны для мелководных зон рифа и особенно для риф-флета, где вода, проходя над рифом во время приливов и отливов, находится в наибольшем контакте с донным населением рифа. Причину убыли фитопланктона на мелководных участках флета следует видеть в его выедании донными фильтраторами рифа (Glynn, 1973а). К тому же клетки днатомей, таких, как Nitzschia delicatissima или ветвистные клетки Chaetoceros, могут травмироваться в турбулентных водах рифа, особенно при столкновениях с твердой поверхностью рифа и с частицами взвеси. В водах же приглубых частей лагуны или за внешним краем рифа, где контакт вод с рифом уменьшается, происходит интенсификация развития фитопланктона за счет отдачи части биогенов в оду донными сообществами рифа.
По количественным показателям его биомассы и продукции, по концентрации хлорофилла фитопланктон в зоне рифа богаче, чем в окружающих водах открытого океана (Michel, 1969; Сорокин, 1971; Ricard, 1976; Sournia, Ricard, 1976а). На рис.6.3 была представлена типичная картина изменения первичной продукции фитопланктона на разрезах от рифа Херон (Б. барьерный риф) в открытый океан. Уже на расстоянии 6-9 км она убывает более чем на порядок величин.
Прозрачность вод по белому диску в приглубых участках рифов обычно достаточно велика и составляет в среднем 10-15 м, хотя в отдельных случаях она может быть меньше (5 м на атолле Кантон) и больше 30 м (атолл Маджуро). Таким образом, фотосинтез фитопланктона практически не лимитируется светом во всей их толще. На рис. 6.11 приводятся типичные кривые ослабления падающей радиации в водах некоторых рифов Индийского океана со средней прозрачностью (10-18). Как видно из этого рисунка, фитопланктон может испытать некоторый недостаток света лишь у самого дна лагуны.
Примеры сезонной изменчивости фитопланктона представлена на рис. 6.9,6.12,6.13. Они показывают, что в разные сезоны года не только меняется его состав (см.6.2.1), но происходят изменения его биомассы и функциональной активности. Колебания величин первичной продукции фитопланктона и концентрации хлорофилла на рифах в течение сезона достигали порядка величин (Angot, 1968; Sournia, 1972). При этом даже в лагуне атолла Такапото, расположенного в районе олиготрофных вод, концентрация хлорофилла в период сезонного максимума достигала уровня, характерного для мезотрофных вод (Sournia, Ricard, 1976). В водах барьерного и берегового рифов его концентрация бьта близка к 1 мг/м 3 , что соответствует его содержанию в евтрофных водах, а биомасса фитопланктона в зоне рифов у о-ва Носи Бе превышала 3 г/м 3 . Значительные сезонные колебания концентрации хлорофилла в воде (от 0,13 до 0,5-2,5 мг/м 3) были отмечены в лагуне Б. барьерного рифа Австралии у г. Таунсвиль (Sammarco, Crenshaw, 1984). Максимум был отмечен в летне-осенние месяцы с декабря по апрель. Он совпадал с периодом дождей и увеличением поверхности стока (рис. 6.14). В этот период в водах лагуны отмечаюсь интенсивное развитие фитопланктона, значительную долю которого составляли синезеленые водоросли Trichodcsmium (Relevante et al., 1982). Стимулирующее влияние речного стока на развитие фитопланктона отмечено и для вод рифов у Новой Каледонии (Binet, 1984). Представленные данные указывают на неосновательность тезиса (Parsons et al., 1977) о том, что планктонные сообщества тропических вод в отличие от умеренных находятся в состоянии некоего гомеостаза и не испытывают существенных сезонных изменений. Такие представления были, по-видимому, следствием недостатка соответствующих сезонных циклов наблюдений в тропических водах (Sournia, 1969). Причина сезонных максимумов в развитии фитопланктона прибрежных тропических вод состоит прежде всего в изменении циркуляции вод на прилегающей к рифам акватории океана, которые имеют годовую периодичность. Причем эти изменения даже в тропической зоне чаще всего имеют место в осенний и весенний период. Не случайно поэтому и сезонные максимумы развития фитопланктона отмечены осенью или весной.
В ходе изучения развития фитопланктона в водах рифов установлено наличие значительных суточных колебаний состава и численности его сообществ, а также его продуктивности (Milliman, Mahnken, 1972; Sournia, Ricard, 1976; Сорокин, 1979). Такие колебания имеют место и в сообществах фитопланктона открытого океана. Они вызываются там суточной периодичностью роста фитопланктона и интенсификацией выедания водорослей ночью зоопланктоном, мигрирующим из нижележащих слоев воды в эвфотическую зону. Те же причины действуют и в прибрежных водах в зоне коралловых рифов, вызывая суточную периодичность развития фитопланктона. Но в этих водах на них накладывается еще влияние суточного приливного цикла. Как показали результаты суточных наблюдений на рифах у островов Намзу, под влиянием выедания мигрирующего ночью в толщу вода фито1шанктона зоопланктоном рифа суточная его продукция ранним утром снижается на порядок величин. Однако уже к 11ч популяция фитопланктона в значительной степени восстанавливается, особенно у края рифа, где фитопланктон интенсивно размножается (рис. 6.15).
Каковы же причины возрастания биомассы и продукции фитотанктона в рифовых водах по сравнению с окружающими водами открытого океана? Вопрос этот достаточно сложен, особенно ввиду того, что такое возрастете имеет место при очень низких абсолютных концентрациях биогенов, мало отличающихся от таковых в открытом океане, а также в условиях необычайно интенсивной эксплуатации сообществ фитопланктона фильтрирующей фауной рифа. Высказанные по данному поводу мнения весьма противоречивы (Sournia, Ricard, 1976; Sournia, 1976б; Salvat 1982). Большая часть авторов в качестве основной причины указывают на так называемый островной эффект ("island mass effect"). Этот эффект заключается в обогащении биогенными элементами вод прибрежной зоны у‘ островов за счет взаимодействия их массы с набегающим потоком океанских течений, в результате чего возникает подъем глубинных вод к поверхности. Эффект обогащения неритической зоны у островов, расположенных в стрежне пассатных течений, таких, как, например, Маркизекие острова (Sournia, 1976в). Б. барьерный риф (Gilmartin, Relevante, 1974), Гавайи (Doty, Capurro, 1959), Сейшельские острова (наши наблюдения), несомненно, имеет место. Под его воздействием первичная продукция в неритической зоне у таких островов бывает достаточно высокой, что влияет на уровень продуктивности и самих рифовых вод. Однако и в этих условиях продукция и биомасса фитопланктона в водах лагун рифов обычно выше, чем в прииегающих к ним и находящихся с ними в состоянии интенсивного обмена водах неритической зоны (Marshall, 1965; Michel, 1969; Sournia, Ricard, 1976, 1976а; Thompson, Golding, 1982). В свете этих данных можно сделать вывод о том, что основная причина повышенной продуктивности фитопланктона в рифовых водах состоит в постоянной отдаче в толщу воды донными и планктонными гетеротрофами биогенных элементов (прежде всего азота, фосфора), которые образуются в результате процессов деструкции органического вещества, протекающих на рифах с большой силой. В то же время ввиду очень высокой скорости оборота биогенов в толще воды рифов далеко не всегда можно зарегистрировать прирост абсолютный концентраций солевых биогенов в водах в зоне рифа по сравнению с прилегающими водами океана (рис. 5.1.).